
La infección producida por el Virus Zika es otra de las arbovirosis transmitidas por mosquitos que han emergido rápidamente a escala global en ambiente urbano en los últimos años. En las Américas, la epidemia de Zika se ha superpuesto sobre otra reciente, la del Virus Chikungunya, lo que se suma a la ya endémica del Virus Dengue, determinando que actualmente circulen en la región 6 infecciones virales transmitidas principalmente por el mosquito Aedes aegypti, sumando los 4 serotipos que circulan normalmente del Dengue. La epidemia, aparentemente originada en Brasil (2015), se ha extendido ya a 40 países/territorios en las Américas y el Caribe infectando a más de 2 millones de personas. Los continuos hallazgos sobre la expresión de este virus y su transmisión ilustran la complejidad eco-epidemiológica de las interacciones virusmosquito. Solo un abordaje multidisciplinario permitirá entender y afrontar esta considerada pandemia por la organización mundial de la salud. En este trabajo, caracterizaremos y describiremos los principales mecanismos de transmisión del Zika, y los procesos que determinan los patrones eco-epidemiológicos de la infección. Igualmente, discutiremos qué factores explican la emergencia de las recientes arbovirosis en el mundo, y que estrategias deberían seguirse a fin de prevenir o mitigar el impacto de ellas. Finalmente, mencionaremos brevemente el estado actual de la epidemia del Virus Zika en Venezuela.
Palabras clave: Arbovirus, Flaviviridae, Zika, Mosquitos, Vector, Aedes aegypti, Aedes albopictus, Venezuela.
Zika virus is up till now another mosquito-borne virus (arbovirus) that is rapidly emerging in an urban setting, on a global scale. In the Americas, it has emerged on the heels of a recent chikungunya epidemic. Therefore, currently, there are 6 viral infections circulating and transmitted by Aedes aegypti in the continent if we include the 4 endemic serotypes of dengue. A Zika epidemic that began in Brazil in 2015 has now spread rapidly to more than 40 countries in the Americas and the Caribbean, infecting more than 2 million inhabitants. The continuous discovery of the ways of transmission and the clinical and epidemiologic expressions of this virus show the complexity of the virus-host-mosquito interactions. This epidemic currently continues unabated. The limited understanding of this virus presents an enormous challenge for responses to this rapidly emerging threat to human health and demands a multidisciplinary approach. This article reviews the existing knowledge about mechanism of transmission, ecoepidemiological patterns and main factors accounting for the emergence of this arbovirus in the world. Also, it discusses ways of vector control and the current status of Zika transmission in Venezuela.
Key words: Arbovirus, Flaviviridae, Zika, Mosquitos, Vector, Aedes aegypti, Aedes albopictus, Venezuela.
Laboratorio de Biología de Vectores y Parásitos, Centro de Ecología y Evolución, Instituto de Zoología y Ecología Tropical, Facultad de Ciencias, Universidad Central de Venezuela.
Caracas, Venezuela.
Las infecciones transmitidas por mosquitos (Diptera: Culicidae) han sido una continua amenaza a la salud humana por décadas. Entre los artrópodos hematófagos (Tabla 1), los mosquitos son los vectores de patógenos más frecuentes, existiendo hasta el momento unas 300 especies perteneciente a los géneros Aedes y Culex involucradas en la transmisión de aproximadamente unos 115 y 105 tipos de Arbovirus, respectivamente (Liang et al. 2015). Una alta proporción de estos patógenos se asocian con enfermedades de gran impacto en humanos y animales (Arbovirosis, Tabla 1), y circulan principalmente en regiones tropicales y subtropicales donde las especies de mosquitos son diversas y abundantes. Entre estas arbovirosis, sin duda alguna, las más comunes y/o de reciente relevancia son las infecciones urbanas del Dengue (VDEN), Fiebre Amarrilla (VFA), Chikungunya (VCHIK) y la más reciente y emergente infección del virus del Zika.
ARBOVIRUS: acrónimo en inglés de “Arthropod-Borne Viruses” definiendo a todos aquellos virus transmitidos biológicamente entre artrópodos hematófagos (ejemplo: mosquitos y garrapatas) y vertebrados, durante el ciclo de vida del virus.
VECTOR: organismo (artrópodo hematófago) que transmite biológicamente un patógeno (virus, parásitos). Los virus o parásitos se replican o desarrollan dentro del vector como parte de su ciclo de vida. Esto es opuesto a ser transmitidos mecánicamente, sin replicación o desarrollo dentro del vector, a través de partes bucales contaminadas.
HEMATOFAGIA: hábito de alimentarse de sangre de vertebrados.
ANAUTOGENIA: requerimiento de ingesta de sangre (como recurso) para el crecimiento, muda, desarrollo de huevos y fitness reproductivo (Ciclo gonadotrófico en insectos). ANTROPOFAGIA/ANTROPOFILIA: preferencia de ingestas sanguíneas sobre hospedador humano, opuesta a zoofagia/zoofilia, hechas sobre hospedadores animales no humanos.
ZOONOSIS: infección viral transmitida entre animales no-humanos.
ANTROPONOSIS: infección viral transmitida de humanos a otros animales. COMPETENCIA VECTORIAL: habilidad intrínseca (o alta susceptibilidad innata) del insecto o artrópodo a permitir el desarrollo completo del patógeno dentro de su cuerpo.
CAPACIDAD VECTORIAL: promedio de casos secundarios de una enfermedad derivados de un primer caso infeccioso en una población susceptible definida. Cuantifica habilidad de la población del vector para transmitir un agente infeccioso. Es una función de la tasa de contacto hombre-vector, la densidad y sobrevivencia del vector, su competencia vectorial y del periodo de incubación extrínseco del patógeno.
SÍNDROME DE GUILLAIN BARRÉ: síndrome donde el sistema inmunitario del organismo ataca parte del sistema nervioso periférico, afectando particularmente a los nervios que controlan los movimientos musculares así como a los que transmiten las sensaciones de dolor, temperatura o tacto. Produce debilidad muscular y pérdida de sensibilidad en las piernas o brazos.
INMUNIDAD GRUPAL (“Herd immunity”): cuando todos en una población humana son vulnerables a la infección de un patógeno nuevo debido a que nunca han estado en contacto previo con él, y no poseen respuesta inmunológica. En esta condición inicial el patógeno se dispersa rápido y lejos. Conforme se va adquiriendo respuesta inmunológica, la onda epidemiológica local del patógeno va desvaneciéndose en la población.
El Virus del Zika (VZIK), es un arbovirus que pertenece a la familia Flaviviridae, género Flavivirus, y está relacionado filogenéticamente con los Virus del Oeste del Nilo (VON), VDEN y VFA. El VZIK fue aislado por primera vez en Macacos (Macaca mulatta) habitando un bosque denominado Zika (en localidad de Entebbe, cerca de Kampala), en Uganda, en el año 1947 (Dick, 1952).
El primer gran brote se detectó en la Isla de Yap, en abril de 2007 donde se observó una enfermedad caracterizada por erupciones cutáneas, conjuntivitis, artralgias, artritis y fiebre. En junio de ese año el CDC (Centro para el Control de Enfermedades), por sus siglas en inglés, confirmó que las muestras de los pacientes contenían ARN de VZIK (Duffy et al. 2009). Posteriormente, se registraron sucesivos brotes en las siguientes islas del Pacifico Sur: Polinesia Francesa en 2013 (Cao-Lormeau et al. 2013), Nueva Caledonia en 2014 (Dupont-Rouzeyrole et al. 2015), Islas Orientales en 2014 (Tognarelli et al. 2015), e Isla de Cook en 2014 (Musso et al. 2104). Se estimó que el brote de la Polinesia Francesa del año 2013 pudo haber afectado aproximadamente a 28.000 personas, cerca del 11% de la población, que buscaron atención médica basada en los signos y síntomas de la fiebre de bajo grado, erupción maculopapular, artralgia, y la conjuntivitis (Musso et al. 2014). Fue durante este brote, que el síndrome de Guillain-Barré (SGB) se asoció por primera vez con VZIK. También se evidenció el riesgo para la transmisión a través de los bancos de sangre, y se detectó el VZIK (o ARN viral) en el semen, saliva y orina, mientras que la transmisión en la Polinesia Francesa se asoció al mosquito Aedes (Stegomyia) polynesiensis (Cao-Lormeau et al. 2013).
Entre 2013-2014, el VZIK entró al continente americano, aún incierto si fue a través de Brasil (Zanluca et al. 2015) o el Caribe (Faria et al. 2016), pero sin ninguna duda, fue desde Brasil donde comenzó como una gran epidemia y se dispersó por toda la América continental e insular. En esta epidemia se observó una asociación entre la infección por VZIK y el SGB, y ese mismo año, en octubre, también se encontró una posible relación con la microcefalia (Campos et al. 2015; Zanluca et al. 2015).
Hasta la Semana epidemiológica Nº 29 (14 julio 2016), la Organización Mundial de la Salud (PAHO/WHO 2016) reportaba transmisión local autóctona del VZIK en 40 países/territorios de las Américas. Esta pandemia, aunada a la anterior y también reciente epidemia del VCHIK demuestran la gran capacidad de adaptación y dispersión de las arbovirosis usando como vectores a los mosquitos, en este caso Aedes (Stegomyia) aegypti (L) y Aedes (Stegomyia) albopictus (Skuse). Aedes aegypti, de hecho, es considerado el principal vector responsable de la transmisión urbana del VZIK, así como lo es de los virus VDEN, VCHIK y VFA en el continente americano (OMS 2016).
El objetivo del presente trabajo es caracterizar y describir los principales mecanismos de transmisión del VZIK, así como los procesos biológicos y ecológicos que determinan los actuales patrones eco-epidemiológicos de este virus en las Américas. De especial interés será discutir qué factores explican la emergencia y/o reemergencia de las recientes arbovirosis en el mundo, y que estrategias deberían seguirse a fin de prevenir o mitigar el impacto de ellas. Finalmente, mencionaremos brevemente, cual es el status epidemiológico actual del VZIK en Venezuela.
El mecanismo de transmisión biológica más común en los Arbovirus es el horizontal, esto es, cuando el virus, en su estrategia de dispersión y sobrevivencia a corto y largo plazo, pasa de la saliva de un vector infectado a un vertebrado hospedador, al este realizar una ingesta de comida sanguínea sobre ese hospedador (Weaver & Reisen, 2010). Este mecanismo involucra la infección del tracto alimentario del vector post ingesta sanguínea infectada, la diseminación del virus por todo el cuerpo del artrópodo para su eventual replicación en las glándulas salivares, seguido por la inyección de saliva infecciosa durante una siguiente ingesta sanguínea. Los Arbovirus circulan naturalmente en ciclos enzoóticos, principalmente en áreas boscosas, entre fauna silvestre y vectores selváticos (Zoonosis, Tabla 1); sin embargo, bajo ciertas circunstancias o cambios eco-epidemiológicos, el hospedador humano entra en este ciclo de manera incidental, al ser picado por un artrópodo infectado. Otro de los mecanismos de transmisión biológica de las arbovirosis es el vertical (denominado venéreo) y se da cuando el virus pasa de una hembra vectora infectada a su progenie (ambos sexos); sin embargo, este es un mecanismo menos común y solo favorecido cuando las condiciones ecológicas son adversas para la replicación del virus.
En el caso de la transmisión del VZIK, se asume que el mecanismo dominante de transmisión en el continente americano ha sido la transmisión vía mosquitos, sin embargo, la relevancia e importancia epidemiológica de otros modos de transmisión se desconocen (Figura 1, modificado de Rodríguez-Morales et al. 2016). Por ejemplo, la transmisión sexual ha sido documentada en áreas donde no existen los mosquitos vectores (Musso et a. 2014), pero la frecuencia y duración del virus en el semen es desconocida. Igualmente, se han reportado casos de transmisión vertical desde la madre a los hijos (Musso et al. 2014) y de transmisión vía transfusión sanguínea (Musso et al. 2014). El mecanismo de transmisión vectorial (Figura 1) consiste en la toma del virus con la ingesta sanguínea del mosquito hematófago sobre el hospedador infectado. El virus infecta el tejido del aparato digestivo del mosquito y se disemina por todo el cuerpo donde el virus se replica durante un periodo denominado periodo de incubación extrínseco, el cual dura aproximadamente entre 7-10 días (Li et al 2012), para luego migrar hasta las glándulas salivares y ser inyectado nuevamente junto a la saliva del mosquito cuando este realiza otra ingesta sanguínea sobre un siguiente hospedador y donde culminara su ciclo de transmisión al llevarse a cabo el periodo de incubación intrínseco.
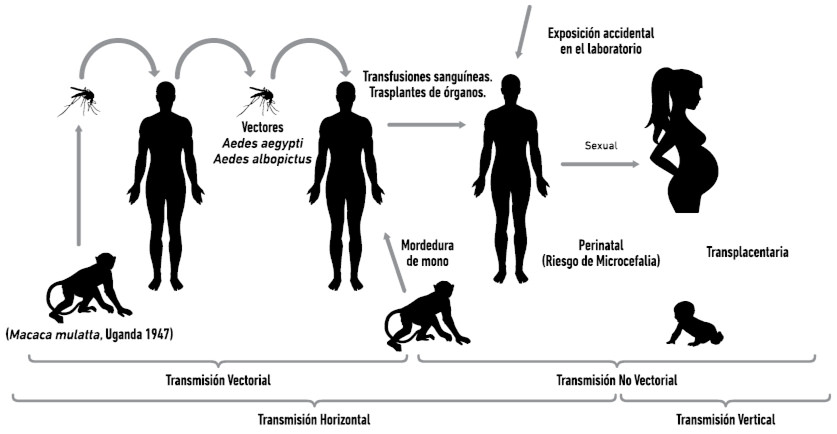
En su medio enzoótico natural en los bosques de África y Asia, el VZIK circula entre especies zoofílicas (Tabla1) de hábitos arbóreos del género Aedes y primates no humanos (Monlun et al. 1993; Kuno et al. 1998). En Uganda, el virus fue aislado por primera vez en un mono hospedador, Macaca mulatta, y en un mosquito vector, Ae. africanus (Haddow et al. 1964). Posteriormente, el virus fue detectado a través de aislamiento viral y de estudios serológicos en humanos infectados, mosquitos y animales en África y Asia (Fagbami 1979; McCrae & Kyria 1982; Akoua-Koffi et al. 2001; Duong et al. 2009; Hayes 2009; Heang et al. 2010) Específicamente, el aislamiento viral fue reportado para los siguientes mosquitos selváticos: Ae.africanus y Ae. apicoargenteus (McCrae & Kyria, 1982), Ae. luteocephalus (Dick et al, 1952; Haddow et al, 1964; Fagbami 1979), Ae. furcifer y Ae. tailori (Monlun et al. 1993), y Ae. vitatus (Diallo et al. 2014). En la epidemia del VZIK, en la Isla de Yap (Micronesia), Ae hensilli fue el mosquito más abundante picando, aunque el virus no fue aislado en esta especie (Duffy et al., 2009). En 2011, en el sureste de Senegal, se registró una amplificación del VZIK (transmisión del virus de no-humanos a humanos), posterior a amplificaciones similares que sufrieron el VCHIK (2009) y el VFA (2010) en la misma región geográfica. El VZIK fue aislado de diversas especies de mosquitos colectados en diversos ecosistemas o paisajes naturales, rurales y urbanos (bosque, plantación agrícola, sabana, comunidades urbanas: extra-domicilio o peri-domicilio) de esa región, sugiriendo la amplia circulación viral observada para ese momento (Diallo et al. 2014). Asumiendo que todos los mosquitos infectados eran capaces de transmitir, estos autores reportaron la siguiente lista de potenciales especies transmisoras del Zika en dicha región: Ae. furcifer, Ae. taylori, Ae.luteocephalus, Ae. vittatus y Ae. africanus. Previamente, Ae. furcifer, Ae. taylori, y Ae. luteocephalus habían sido incriminados como vectores selváticos de VCHKV, VDEN-2 y VFA en la misma área (Cornet et al. 1979; Diallo et al. 2003; Diallo et al. 2012). Cabe destacar varias observaciones y hallazgos del estudio de Diallo et al, (2014). Primero, aunque Ae. aegypti fue colectado infectado, esta especie predominaba solo en el bosque; es decir, era una población selvática. Segundo, Ae. vittatus fue la especie de mosquito sugerida como especie “puente” o especie exportadora del virus desde el ambiente selvático al ambiente doméstico, ya que ella fue la única colectada picando a humanos a todo lo largo de este gradiente espacial: dosel del bosque, sotobosque y peri-domicilio (alrededor de las casas). Finalmente, la especie Ae. furcifer fue encontrada infectada en muestras de machos y no hembras en el ambiente cerca de las casas, sugiriendo la transmisión vertical (o venérea) del virus como un mecanismo de mantenimiento de la circulación local del patógeno en la región. Podemos decir entonces que previo a su reciente expansión en América, el virus Zika predominaba principalmente en ambientes selváticos, aunque ya había sido aislado en ambientes urbanos de África y Asia, en humanos y en el mosquito Ae. aegypti (Fagbami, 1979; Olson et al. 1981; Diallo et al. 2014). De hecho, las primeras implicaciones de Ae. aegypti en la transmisión urbana del virus Zika provienen de evidencias de campo en Nigeria (Fagbami, 1979), las coocurrencias espaciales de picos de infección en humanos con picos de densidad poblacional de Ae. aegypti en Indonesia (Olson et al. 1981) y finalmente, el aislamiento viral en “pooles” del mosquito en Malasia (Marchette et al. 1969), siendo en este último trabajo donde se comprobaba por primera vez la circulación del VZIK fuera de África. Igualmente, estudios experimentales tempranos en Nigeria, Senegal y Singapur habían confirmado la competencia vectorial (Tabla 1) de poblaciones de Ae. aegypti a la infección exitosa con el virus en el laboratorio (Boorman & Porterfield 1956; Marchette et al. 1969; Cornett et al. 1979; Li et al. 2012). Sin embargo, aunque Ae. aegypti se ha mostrado susceptible a la infección oral con el Virus Zika en África, la mayoría de estos experimentos han demostrado una muy baja proporción del genoma viral en la saliva de los mosquitos experimentales, lo que concuerda con las bajas tasas de aislamiento que muestra el virus en poblaciones naturales de este mosquito, e indican una baja capacidad de la especie para diseminar y transmitir el virus en dicha región (Diagne et al. 2015). Adicionalmente, estos hallazgos subrayan la determinación y variabilidad genética de la competencia vectorial en mosquitos (Tesh et al. 1976; Failloux et al. 2002) y el papel del genotipo del patógeno en la interacción virus-mosquito (Fansiri et al. 2013). Es importante mencionar que tres son los linajes del VZIK circulando actualmente en el mundo: Africano-Oriental, Africano-Occidental y Asiático (Faye et al. 2014), a lo cual volveremos más adelante en este texto.
Aedes aegypti, el mosquito antropofílico (Tabla 1) urbano por excelencia (comúnmente llamado mosquito patas blancas) es considerado el primer vector responsable de la transmisión urbana del VZIK en las Américas, así como es el principal transmisor de los VDEN, VCHIK y VFA urbana en la región. Por su parte, se ha demostrado que es el genotipo asiático del VZIK el que invadió y circula actualmente en el continente americano (Faye et al. 2014; Haddow et al. 2012). Este mosquito es originario de los bosques de África Occidental, en donde su forma ancestral desarrolla sus fases inmaduras en huecos de árboles y las hembras tienen hábitos zoofílicos (Tabla 1). Hipotéticamente, fue introducido en el continente americano a principios del siglo XVII, en la época de la colonia, en los barcos que transportaban esclavos (Tabachnick, 1991; Eisen & Moore, 2013). La capacidad que tienen las formas inmaduras (huevos) de muchas especies de Aedes a resistir la desecación por largos periodos de tiempo, los hace ser especies altamente dispersoras e invasoras, pudiendo sus huevos ser transportadas a grandes distancias en barcos y/o aviones (Reiter, 1987). En el continente americano, la ecología de Ae. aegypti cambio, adaptándose al ambiente doméstico humano, a través de la colonización de depósitos acuáticos naturales o artificiales presentes en el área urbana-doméstica, reminiscentes de su hábitat original (huecos de árboles), y haciéndose preferentemente antropofílicos. El papel de Ae. aegypti como transmisor de patógenos causantes de enfermedades al hombre, fue demostrado en 1901 por el médico estadounidense Walter Reed, confirmando la hipótesis planteada originalmente por el médico cubano Carlos Finlay sobre el papel de esta especie en la transmisión del VFA (Cristorphers 1960).
El menos antropofílico y urbano, pero sí muy agresivo mosquito picando, Aedes albopictus (conocido como el mosquito tigre), es considerado el segundo vector del VZIK en las Américas. De hecho, Ae. albopictus ha sido encontrado infectado naturalmente con VZIK en el viejo continente (Grard et al., 2014) y ha probado ser un vector experimentalmente competente para este virus (Wong et al.; 2013). Esta especie ha expandido su rango de distribución en los últimos 40 años, de un ambiente rural nativo tropical en el sur-este asiático hasta ambientes rurales, semi-urbanos y urbanos del continente americano, presumiblemente a través del transporte de sus huevos resistentes a la desecación, vía el comercio internacional de llantas usadas (Reiter 1987; Benedict et al. 2007). Un elemento clave que ha favorecido la expansión del mosquito Ae. albopictus al ambiente urbano ha sido su habilidad para cambiar de estrategia de colonización de hábitats acuáticos naturales conocidos como fitotelmatas (esto es, huecos de árboles, tallos de bambú, y bromélias, entre otros) a contenedores artificiales tales como, envases abandonados, llantas, floreros de cementerios, y recipientes abandonados que retienen agua de lluvia (Hawley, 1988). De hecho, Ae. albopictus está considerada como una de las 100 especies invasoras más peligrosas para la salud humana, como consta en el catálogo de especies invasoras mundiales (Global Invasive Species Database: http://www.issg.org/database/species). Paralelo a su invasión al nuevo continente, Ae. albopictus ha desplazado o coexiste con Ae. aegypti, su especie competidora en el ambiente urbano, aumentando así su amenaza como vector adicional de las arbovirosis urbanas. Aedes albopictus puede actuar como vector secundario del VDEN (Lambrechts et al. 2010) y ha sido implicado en la transmisión de al menos 26 arbovirosis en el laboratorio (Gratz, 2004); igualmente, se ha confirmado su papel como vector principal del reciente brote del VCHIK en el mundo (Papuy et al. 2009; Vega-Rua et al. 2014).
¿Qué evidencias existen acerca de la transmisión del VZIK por parte de Ae. aegypti y Ae. albopictus en las Américas? Un estudio reciente demostró que cuando ambas especies (poblaciones americanas) eran infectadas experimentalmente en el laboratorio con las cepas del virus que circula actualmente en el continente (genotipo asiático), aunque la infección se diseminaba en el cuerpo del mosquito, la proporción del virus en saliva era muy baja (Chouin-Carneiro et al. 2016), indicando una baja competencia vectorial (Tabla 1) de Ae. aegypti y Ae. albopictus por la transmisión del VZIK. Estos resultados sorprenden y requieren ser profundizados, complicando el rompecabezas de la transmisión del VZIK en las Américas, más aun, cuando este virus se ha diseminado de manera muy rápida en el continente desde su llegada. Sin embargo, y asumiendo que la vía vectorial es el principal mecanismo de transmisión, solo una alta densidad vectorial, principalmente de Ae. aegypti, que condicionaría una alta tasa de contacto hombre-vector (principalmente en el domicilio), es lo que explicaría en parte la alta transmisión del VZIK en Latinoamérica, a pesar de la baja competencia experimental observada. Por supuesto, otros factores atribuibles al hospedador humano deben estar jugando un papel importante en esta rápida diseminación del virus, como lo es el hecho de ser un nuevo patógeno interactuando con una población hospedadora inmunológicamente nueva y altamente susceptible a la infección. Sin embargo, considerando nuevamente al vector, de lo que sí existe evidencia en el continente, es de las altas abundancias y amplia distribución en el ambiente urbano del mosquito Ae. aegypti, producto del fracaso de las medidas de control y manejo de esta especie en los últimos 30 años (Powell & Tabachnick, 2013). Es bien conocido que Ae. aegypti es el principal responsable de la transmisión del VDEN y VFA en toda Latinoamérica, y que la infestación generalizada del mosquito en la mayor parte de las ciudades de nuestros países ha sido la causa de las recientes epidemias ocasionadas por VCHIK en 2013-2014. Por otro lado, es bueno ilustrar que poblaciones de Ae. aegypti con baja competencia vectorial pero altas densidades de picada han estado ligadas a brotes de VFA en Nigeria (Miller et al. 1989), enfatizando la importancia de considerar los distintos componentes que forman parte de la capacidad vectorial (Tabla 1) de una especie vectora e ilustrando la complejidad eco-epidemiológica de las interacciones virus-mosquito.
Es un virus de ARN, envuelto, con cápside icosaedrica, no segmentado, monocatenario positivo, que contiene 10794 nucleótidos que codifican 3419 aminoácidos. Está estrechamente relacionado con el virus Spondweni; siendo estos dos virus los únicos miembros de su clado dentro de la agrupación de los Flavivirus (Lanciotti et al, 2008; Kuno et al 1998; Cook & Holmes, 2006). Los grupos más próximos incluyen a los virus Ilheus, Rocío, San Luis, y Fiebre Amarilla (que es el prototipo de la familia); pero también incluye al Dengue, la Encefalitis Japonesa, y el virus del Nilo Occidental (Lanciotti et al, 2008, Kuno et al. 1998).
Existen tres linajes de VZIK, dos africanos (Africana-Oriental, Africana-Occidental) y uno asiático (Asiática), y estudios filogenéticos indican que el virus que está expandiéndose por el continente americano está más estrechamente relacionado con cepas provenientes de la Polinesia Francesa o con el linaje asiático (Fayes et al. 2014). En este trabajo, las cepas o linajes africanos pueden clasificarse en dos grupos, un clado en donde se ubica la cepa prototipo (Uganda) y un clado donde se ubican las muestras de Nigeria; las cepas correspondientes a Micronesia y Malasia forman otro clado y es el llamado linaje Asiático. Las cepas de Costa de Marfil y Senegal en ambos grupos africanos, sugieren que al menos dos linajes distintos de ZIKV circularon en estos países. Esto autores también sugieren dos introducciones del VZIK independientes en África occidental desde la parte oriental del continente. Una primera introducción viral en Costa de Marfil y Senegal que se relacionó con el clado de la cepa prototipo, indicando que posiblemente el virus se dispersó desde Uganda a Senegal en 1985 y a Costa de Marfil en 1995, aproximadamente. La segunda introducción se relaciona con la de Nigeria, cuando el VZIK probablemente se dispersó desde Uganda a la República Centroafricana y Nigeria. Por otra parte, un linaje ZIKV adicional de Uganda probablemente se extendió a Malasia alrededor de 1945 y desde allí, el virus llegó a Micronesia en 1960, formando el grupo de Asia. Por lo tanto, suponiendo un origen del VZIK en Uganda, estos resultados revelaron al menos dos salidas independientes de África Oriental de acuerdo con los dos clados africanos propuestos anteriormente y también apuntaron a un flujo migratorio viral de África oriental a Asia.
Weaver et al. 2016, realizaron análisis con la secuencia completa de VZIK, obteniendo resultados concordantes con los de Fayes et al. 2014, reiterando que es factible la hipótesis del origen africano del virus y posterior divergencia en dos grandes linajes principales: linaje Áfricano y linaje Asiático /Americano. Las cepas africanas se dividen a su vez en dos grupos distintos. El primer grupo (Uganda) está anclado por la cepa prototipo e incluye aislamientos de Senegal y la República Centroafricana 1947-2001; mientras que el segundo grupo (Nigeria), incluye cepas aisladas de Nigeria. Finalmente, las muestras provenientes de la epidemia Americana se pueden asociar o incluir en el linaje Asiático (Micronesia, Malasia).
Es en el medio acuático donde se llevan a cabo los eventos de crecimiento y desarrollo de las fases inmaduras (pre-adultos) de las especies de Aedes, por lo que la escogencia del sitio donde colocar los huevos (ovoposición) por parte de una hembra grávida es un rasgo de la historia de vida del mosquito de importancia crítica y de importancia epidemiológica para la transmisión de arbovirus. Adicionalmente, la ubicación, abundancia, y diversidad de hábitats de inmaduros serán los principales determinantes de la distribución y abundancia espacial y temporal de las poblaciones adultas de mosquitos. De allí que identificar estos sitios es una tarea básica para conocer los hábitats potenciales donde larvas y pupas eventualmente serán encontradas y donde los mosquitos adultos emergerán, a fin de diseñar estrategias de vigilancia y manejo más efectivas. Los principales hábitats colonizados por las hembras de Ae. aegypti en áreas urbanas son todos aquellos recipientes abandonados o no que acumulan agua de lluvia o agua depositada tales como las piscinas o fuentes, los tanques de almacenamiento de agua para consumo, las llantas o cauchos usados descartados, los floreros de cementerio, las latas y botellas descartados en la basura, los canales de desagüe en los techos y sumideros de agua, las cisternas, y los hábitats naturales (ejemplo: plantas) que retienen agua. Generalmente, Ae. aegypti coloniza hábitats con agua limpia, sin embargo, su estrategia puede estarse diversificando al colonizar nuevos hábitats, como lo que se ha observado en Puerto Rico, en donde los pozos sépticos con alto contenido de materia orgánica en descomposición producen cantidades importantes de esta especie (Barrera et al. 2008). El control vectorial puede hacerse menos eficiente donde está presente Ae. albopictus, ya que esta especie no solo coloniza habitas acuáticos similares a los de Ae. aegypti, sino que también coloniza hábitats naturales normalmente no considerados en los programas o medidas de control de vectores de estas arbovirosis. Por otra parte, un factor importante a considerar es la falta del servicio continuo de suministro de agua potable en las ciudades y barrios más pobres de América Latina y el Caribe lo cual obliga a las familias a almacenar agua en tanques, y baldes, entre otros. Consecuentemente, estos contenedores se convierten en el mejor sitio de reproducción del mosquito dentro y alrededor de las casas, aumentando el riesgo de incrementar las densidades de adultos picando, lo que aumenta la tasa contacto hombre-vector y el riesgo de contraer la infección viral. No hay que olvidar que Ae. aegypti es una especie altamente antropofilica, de allí que este mosquito puede coexistir con nosotros dentro de las casas si encuentra las condiciones apropiadas para su desarrollo y reproducción. De hecho, la transmisión de una arbovirosis como el VDEN ocurre de manera muy focalizada dentro o alrededor de las casas, debido a que Ae. aegypti no vuela más allá de los 100 metros desde sus sitios de cría y ovoposición. Por lo tanto, la presencia de los hábitats acuáticos donde se desarrollan los inmaduros de Ae. aegypti o Ae. albopictus dentro o alrededor de las casas es el principal factor de riesgo epidemiológico de contraer el VZIK en área urbana.
Las Zoonosis (Tabla 1) suelen desarrollarse en ecosistemas naturales, rurales o suburbanos en complejas interacciones eco-epidemiológicas que involucran a varias especies vectoras y hospedadores vertebrados. Para que una amplificación viral progrese rápidamente y pase de un nivel de circulación enzoótico a niveles epidémicos (transmisión del virus entre humanos y mosquitos) se requiere que al menos exista un vector competente (mosquito) que interactúe (entre en contacto) frecuentemente, en un ambiente ecológico apropiado, con poblaciones hospedadoras de vertebrados susceptibles. Los humanos se exponen a las Zoonosis cuando invaden sus ambientes naturales o cuando un mosquito vector actúa como especie “puente”, sacando el virus de su ciclo enzoótico e incorporándolo a un ciclo epizoótico (virus circula entre mosquitos-vertebrados-humanos) en un ambiente rural o semi-urbano (Weaver & Reisen, 2010). Normalmente, las arbovirosis persisten en baja frecuencia, circulando de manera “silente” para el humano, en sus ciclos naturales (ejemplo: bosque), hasta que bruscamente, surgen cambios genéticos en el virus, cambios en la dinámica poblacional de las especies vectores y/o hospedadores, cambios en la composición de especies vectoras o cambios ambientales naturales o antropogénicos que facilitan la rápida amplificación, dispersión a larga distancia y finalmente invasión del virus a nuevos paisajes epidemiológicos (Weaver & Reisen, 2010).
Un primer factor a considerar cuando se observan brotes inusuales epidémicos (emergentes) es el cambio genético del virus, el cual pudiera o bien incrementar su virulencia en el hospedador vertebrado, incrementando su amplificación potencial y/o aumentar la competencia vectorial y capacidad de transmisión de las especies de mosquitos. Esta hipótesis ha sido explorada en el caso del VZIK comparando las cepas que circulan actualmente en las Américas con las cepas de África y Asia y su capacidad de infectar al mosquito vector. Sin embargo, estos estudios no han confirmado que la cepa del VZIK que circula en las Américas sea más virulenta que las cepas originales del viejo continente, ni que el vector sea más competente y mejor transmisor (Datos citados y aun no publicados en Maron 2016).
Pasamos entonces a considerar factores exógenos y allí los cambios en el uso de la tierra tales como la deforestación y la urbanización pueden jugar un papel muy importante al aumentar el nicho y la abundancia de especies vectoras favorecidas por el aumento de la vegetación de borde (ecotonos) que se forma en estos casos, y actuando estas como las especies “puente” entre el ciclo enzoótico y epizoótico del virus. Ya mencionamos anteriormente el papel jugado por la especie Ae. vittatus en los bosques de Senegal (Diallo et al, 2014), al ser la exportadora del VZIK del ambiente selvático al ambiente doméstico, ya que tiene la capacidad de alimentarse a todo lo largo del gradiente espacial, esto es, dentro del bosque, en el sotobosque y dosel del bosque, y fuera del bosque, en el peri-domicilio (alrededor de las casas). De hecho, María Eugenia Grillet, Fabiola Del Ventura estos mismos autores han evidenciado como la deforestación por la minería, el cultivo agrícola y la urbanización ha sido la principal causa de la degradación de los bosques de Senegal y posiblemente han contribuido con el aumento de la ocurrencia y de las densidades de especies como Ae. vittatus (Diallo et al, 2014).
Sin duda alguna, la globalización y el incremento significativo del comercio e intercambio humano a nivel internacional ha sido otra de las causas principales de la invasión de nuevos patógenos y vectores a nuevas latitudes y continentes. Los ejemplos de invasión de los vectores Ae. aegypti y Ae albopictus al nuevo continente ya fueron mencionados en este trabajo previamente. Es importante mencionar como la reciente invasión de Ae. albopictus al continente americano ha sido seguida por la invasión de los VCHIK y VZIK que el transmite. Las evidencias de cómo primero el VCHIK y ahora el VZIK ha “saltado” desde África y Asia a el continente americano, ilustran el papel significativo que ha tenido la alta interconexión aérea y fluvial que existe actualmente en el mundo, lo que permite que los hospedadores infectados puedan diseminar el virus de manera muy rápida contribuyendo a las grandes epidemias.
Finalmente, la receptividad del paisaje epidemiológico en una región geográfica determinada a la invasión de un patógeno se relaciona con todas aquellas condiciones ecológicas y sociales adecuadas a la replicación y amplificación exitosa del virus, como lo serían: un clima adecuado (altas temperaturas, precipitación y humedad), vector(es) susceptible(s) y abundante(s), vector(es) altamente antropofílico(s), hospedador susceptible debido a la ausencia de inmunidad grupal (Tabla 1) o de la horda (“Herd inmmunity”), condiciones socioeconómicas que favorezcan la expansión del vector y la transmisión, tales como el crecimiento desordenado urbano (pobreza), la deficiencia en el suministro de agua (ya mencionado) y las “islas de calor” que representan los centros urbanos facilitando la transmisión del virus (Weaver & Reisen, 2010). Sin duda alguna, todos estos factores actuando a distintas escalas espaciales y temporales han jugado un papel importante, en un menor o mayor grado, para la invasión exitosa del VZIK en las Américas.
Aunque hemos mencionado que existen factores multifactoriales que explican el aumento y expansión explosiva del VZIK en Latinoamérica, una de las causas primordiales ha sido que el virus invadió centros urbanos donde los niveles del principal mosquito vector, Ae. aegypti, eran o son alarmantes, producto de la existencia de programas de control ineficientes o inadecuados. Las malas o inexistente políticas de salud pública, la resistencia de los mosquitos a los insecticidas en uso, la falta de personal capacitado o entrenado, entre otras, caracterizan hoy en día a muchos de los programas de control en Latinoamérica. Sin embargo, mientras no se cuente con una vacuna, el control y manejo integrado del vector (MIV) es la única opción disponible y adecuada para combatir al VZIK, al igual que ha sido y debería ser la estrategia para combatir al VDEN y VCHIK.
Se debe empezar por mejorar la vigilancia epidemiológica continua y sostenida, a fin de que esta pueda ser una herramienta incluso de prevención o alerta temprana. Igualmente, se debe mejorar el manejo de casos de la infección al fin de contar con técnicas de diagnóstico prácticas, que permitan el reporte clínico efectivo o veraz en los centros de salud, tanto públicos como privados. El desarrollo, mantenimiento y modernización de los sistemas de información epidemiológica del país son tareas urgentes a fin de orientar al personal de salud y porque no, a la comunidad, en donde y cuando se están produciendo los casos sintomáticos del virus. Acá juegan un papel importante las nuevas herramientas tecnológicas tales como la telefonía celular y las redes sociales. Esto permitiría a los organismos competentes elaborar mapas epidemiológicos a distintas escalas espaciales (manzanas, vecindarios, sectores, alcaldías, parroquias, municipios y estados) y temporales que priorizarían las labores de control, enfatizando el control integral de los focos calientes o áreas más activas de transmisión del virus. Hay necesidad de actualizar los métodos de vigilancia efectiva de las poblaciones Ae. aegypti y ahora de Ae. albopictus en las áreas de transmisión. Se deben evaluar nuevos métodos de monitoreo de las poblaciones del vector, porque los métodos clásicos han resultado ser poco efectivos.
Con el control del vector, los objetivos son reducir la población del mosquito a niveles por debajo del cual se evite la transmisión epidémica del virus e interrumpir lo más que se pueda el contacto hombrevector a fin de reducir significativamente el riesgo de contraer la infección. Evitar promover los sitios de reproducción y cría de los mosquitos en los hogares es una medida que puede ser llevada a cabo por la comunidad con una adecuada campaña educativa y de orientación por parte de sectores gubernamentales y no gubernamentales. Si es inevitable acumular agua en las casas, que esto se haga de manera que se impida la colocación de huevos por parte de los mosquitos en dichos envases de almacenamiento. Igualmente, jornadas de recolección de la basura, saneamiento ambiental y jornadas de fumigación aérea (o aerosol) cerca de las viviendas y obligatoriamente, dentro de ellas, permiten combatir las poblaciones de mosquitos adultas. Acá es importante el uso adecuado de insecticidas aprobados por las leyes de regulación internacional, así como el uso y rotación de los mismos a fin de manejar a resistencia del mosquito a los insecticidas. El uso de agentes químicos, biológicos o ambos en los sitios de cría (acuáticos) por parte de las autoridades locales de salud evitaría la emergencia de adultos picando en y alrededor de las casas. La protección personal dentro de las casas es una medida demandante en áreas de intensa transmisión y abundancia del mosquito. La utilización de mosquiteros durante el día (horas de actividad de los Aedes), el uso de telas metálicas en puertas y ventanas, el uso de aire acondicionado en los espacios residenciales y el uso de repelentes constituyen algunas de estas medidas domésticas.
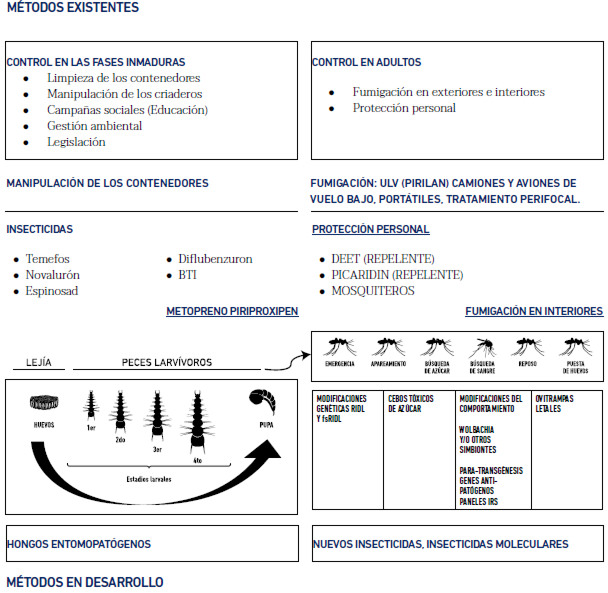
En la Figura 2 (modificada de Achee et al. 2015) se resumen algunos de los métodos existentes y/o en desarrollo de control de Aedes, tomando en cuenta si son de tipo químico, biológico, o físico (remoción); si son de uso personal, comunitario o llevado a cabo por el personal de salud; y sobre que fases del mosquito actúan. Tecnologías de control de mosquito novedosas están actualmente bajo prueba experimental, entre ellas cabría destacar la producción y liberación, en el sitio de transmisión, de mosquitos modificados genéticamente o bien infectados con algún agente (ejemplo: bacteria Wolbachia) que bien impida la reproducción efectiva de los Aedes (Carvalho et al. 2015) o bien disminuya la competencia vectorial del mosquito (Aliota et al. 2016). La idea subyacente es que estas poblaciones de mosquitos modificados genéticamente, estériles o refractarios a la infección, al ser liberados, deberían acabar o reemplazar en el tiempo y en el espacio a las poblaciones naturales de Ae. aegypti. Solo el tiempo dirá si estas técnicas puedan ser efectivas en el control e interrupción de la transmisión local no solo del VZIK, sino del VDEN y VCHIK. Finalmente, es importante recalcar que evitar o combatir a estas arbovirosis es un compromiso de todos. Por otro lado, las medidas de control por parte de los organismos de salud requieren que ellas sean implementadas de manera sostenida, respetando las características locales de cada área a controlar y con coberturas máximas (> 80 %) que alcancen de manera efectiva a la población bajo riesgo.
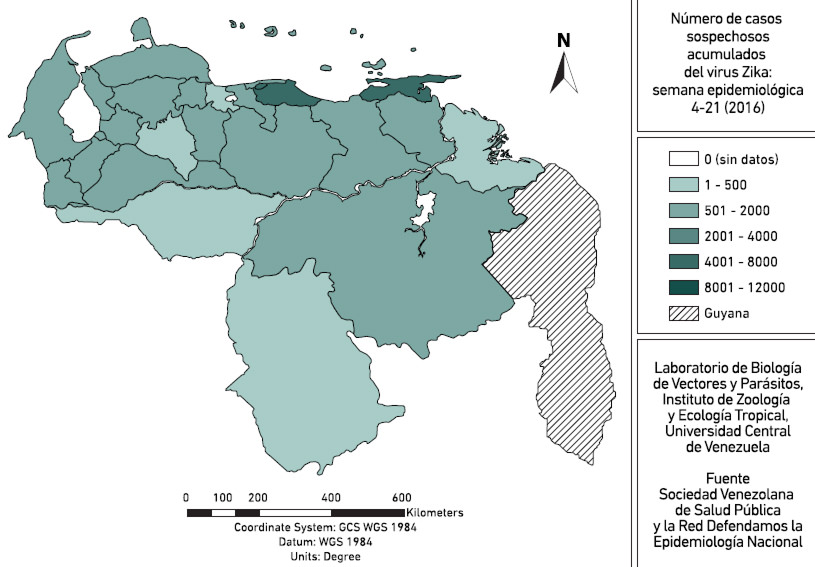
Hasta la Semana epidemiológica Nº 21 (2016), la Sociedad Venezolana de Salud Pública y la ONG Red Defendamos la Epidemiologia Nacional contabilizaba un total de 37558 casos sospechosos acumulados del VZIK en Venezuela durante 2016 (Figura 3), con la mayor incidencia de casos en los estados Distrito Capital, Miranda y Sucre. Sin embargo, de acuerdo a esta ONG, desde junio/julio de 2015 hasta la fecha se podrían haber presentado más de 500.000 casos de Zika en el país, tomando en cuenta que la información epidemiológica no ha sido registrada de manera efectiva por parte del MPPS y que la vigilancia de casos no se ha llevado a cabo de manera activa y obligatoria. Para la S45 del año 2015, los índices aédicos, que son los que cuantifican aquellas casas positivas a la presencia de Ae. aegypti en el país, de acuerdo al Boletín Epidemiológico del MPPS, eran mayores al 5% en casi todo el país hasta llegar en algunos estados norcentrales a alcanzar valores > 20%. La OMS recomienda índices menores del 5% como umbral adecuado de densidades de mosquitos en área endémica para evitar ciclos epidémicos del virus. Estos altos niveles de densidad vectorial se suman al hecho de que nada sabemos acerca de las densidades actuales de Ae. albopictus, especie invasora detectada por primera vez en 2009 en el país (Navarro et al. 2009), todo lo cual nos hace alertar y llamar la atención acerca de la importancia y urgencia de activar la vigilancia epidemiológica, incluyendo la entomológica, y retomar los actualmente inexistentes programas de control de los mosquitos transmisores del VZIK, VDEN y VCHIK en Venezuela.
Las autoras agradecen a la APIU la invitación a participar y contribuir con esta revisión en este número especial sobre la transmisión y emergencia del virus Zika en Latinoamérica. Agradecemos al Dr. José Oletta, a la Sociedad Venezolana de Salud Pública y la ONG Red Defendamos la Epidemiologia Nacional por suministrarnos la data de casos sospechosos de Zika en Venezuela (2016). Finalmente, agradecemos al Lic. Juan Vicente Hernández Villena quien gentilmente realizo el mapa de esta publicación.