Introducción
Entre las emergencias virales del siglo XXI, varias han tenido una resonancia particular en el continente americano. Entre ellas, cabe mencionar la pandemia de influenza H1N1pd en 2009 y la triple epidemia actual de dengue, chikungunya y más recientemente Zika en la región. Esta revisión pretende cubrir la biología de este virus, así como intentar explicar las razones de esta emergencia, la inmunopatología comparada de estos virus y proponer algunas medidas de control de este grave flagelo.
Virus Emergentes
La definición amplia de virus emergentes (Pujol, 2000) comprende a:
- Virus cuya incidencia ha aumentado significativamente en las últimas décadas en una región geográfica determinada, como ha ocurrido recientemente y con cierta frecuencia con algunos arbovirus (virus transmitidos por artrópodos), debido principalmente a fenómenos climáticos y al incremento de poblaciones de mosquitos del género Aedes (Weaver et al. 2010; Chadee et al. 2016). La Figura 1 muestra la emergencia de virus o cepas virales en el Continente Americano, con grandes implicaciones en Salud Pública. Otro ejemplo de este tipo de emergencia lo constituyen las pandemias de influenza. Las pandemias ocurren generalmente por el advenimiento de variantes, producto del rearreglo génico de sus segmentos genómicos. Al ser estos virus de genomas segmentados, se generan nuevos virus con características antigénicas y tropismo particular hacia el humano, que pueden provocar epidemias de gran magnitud (Trombetta et al. 2015).
- Virus recientemente descubiertos, gracias al desarrollo de herramientas cada vez más poderosas de biología molecular. Los virus causantes de hepatitis han sido un ejemplo de virus emergentes.
- Virus realmente nuevos, como lo fue el VIH al inicio de la epidemia. Más recientemente, podemos mencionar al virus responsable del Sindrome Respiratorio Agudo Grave o SRAG (SARS, por sus siglas en ingles), perteneciente a la familia coronavirus (Su et al. 2016). Estos virus a menudo son productos de saltos de especie de un virus animal, que sufre cambios significativos para generar un nuevo virus con capacidad de infectar a humanos.
El virus Zika pertenece a la primera categoría por su emergencia explosiva en las Américas. Las razones de emergencias sucesivas de arbovirus en las Américas se deben a distintos factores:
- El alto número de individuos susceptibles a la infección por el virus. Al ser un virus nuevo en el continente y no existiendo evidencia de protección heteróloga por otros flavivirus, como dengue o fiebre amarilla, virtualmente el 100% de la población americana es susceptible a la infección por virus Zika (VZIK).
- El incremento en infestación de mosquitos, tanto de Aedes aegypti como de Aedes albopictus, ambos vectores responsables de la transmision del dengue, chikungunya y Zika (Tolle, 2009; Weaver et al. 2010). El cambio climático ha sido un factor determinante en el incremento de poblaciones de mosquitos en la región (Chadee et al. 2016).
- La adaptación de estos virus a mosquitos urbanos (Musso et al. 2016).
- La competencia vectorial. No todas las variedades de mosquitos son capaces de infectarse y de transmitir con la misma eficiencia a un arbovirus determinado y estos aspectos determinan la dinámica de la epidemia en una región geográfica determinada (Althouse et al. 2015; Weaver et al. 2010). Sorprendentemente, las evidencias obtenidas hasta la fecha sugieren que tanto Aedes aegypti como Aedes albopictus no parecen mostrar una alta competencia vectorial para el virus Zika (Chouin-Carneiro et al. 2016), no habiendo detenido esto la incidencia explosiva que tuvo la epidemia en la región.
- La rápida evolución del virus Zika, que además de ser un virus ARN, tiene una alta frecuencia de recombinación (Faye et al. 2014), lo cual incrementa enormemente su potencial de adaptación a distintos hospederos vertebrados e invertebrados.
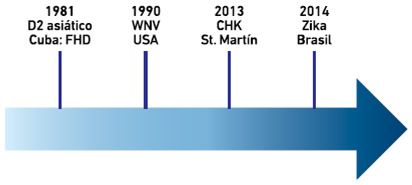
Figura 1: Ejemplos emblemáticos de emergencias de arbovirus en las Américas en los últimos 30 años.
Interesantemente, las introducciones recientes de serotipos o genotipos nuevos de virus dengue (VD) o del virus chikungunya (VCHIK) en Suramérica se han producido desde el Caribe, mientras que la entrada del VZIK a las Américas fue directamente por tierra continental, en Brasil. Esta diferencia puede haber facilitado la rápida diseminación del virus en el continente.
Historia de la emergencia del Vzik
El VZIK es un virus del género Flavivirus, perteneciente a la familia Flaviviridae. El VZIK fue aislado por primera vez en 1947 en el bosque de Zika, en Uganda (África oriental), a partir de la sangre de un mono Rhesus (Macaca mulatta) utilizado como centinela en actividades de vigilancia epidemiológica para el virus de la fiebre amarilla. El año siguiente el virus fue aislado en la misma localidad de mosquitos (Aedes africanus) (Dick et al. 1952). El VZIK fue detectado por primera vez en humanos, de un caso sintomático (fiebre y malestar general) en Nigeria en 1954 (Mac-Namara et al. 1954). Como los virus del dengue y chikungunya, el VZIK es transmitido por artropodos (arbovirus). Como otros arbovirus el VZIK probablemente se adaptó de un ciclo ancestral de transmisión que incluye primates no humanos y como vectores especies de mosquitos arborícolas selváticos, a un ciclo urbano-periurbano con humanos como reservorio y mosquitos Aedes (Stegomya) como vectores (Musso et al. 2015a),
La primera evidencia de circulación del agente en Asia se dio en 1966, aislándose el virus por primera vez a partir de mosquitos urbanos (Aedes aegypti), en Malasia (Marchette et al. 1969). En los años siguientes el virus estuvo asociado a brotes muy pequeños y esporádicos en Asia y en África hasta que en el 2007 hubo una epidemia importante en la Isla de Yap (Micronesia), donde cerca del 75% de la población resultó infectada, con síntomas de fiebre, rash, conjuntivitis y artralgia (Duffy et al. 2009). Evidencias basadas principalmente en datos de serología sugieren que el VZIK es endémico en toda África (excepto en el Norte del continente) y en varios países de Asia.
La siguiente epidemia se dio en la Polinesia Francesa en el Océano Pacifico, en los años 2013-2014, con alrededor de 30.000 casos sintomáticos (Cao-Lormeau et al. 2013). Antes de la epidemia en la Polinesia francesa la infección por el VZIK era normalmente descrita como una enfermedad febril no severa con síntomas que incluían dolores articulares y musculares, rash maculopapular, cefaleas y un aumento muy significativo de casos del síndrome de Guillain-Barré (SGB), una enfermedad neurológica severa, en su forma más grave causante hasta de parálisis. Otras enfermedades causadas por arbovirus como la fiebre del Nilo Occidental, encefalitis Japonesa, chikungunya, y dengue han sido reportadas también como causa de este síndrome (Leis et al. 2012; Ravi et al. 1994; Lebrun et al. 2009; Solomon et al. 2000). Aunque el aumento de número de casos observados fue importante, la circulación concomitante de dengue, que también había sido asociado con el síndrome, no había permitido establecer una relación causal entre la infección por el VZIK y el SGB.
Introducción en las Américas
Hasta el 3 de marzo del 2014, cuando Chile notificó la confirmación de un caso de transmisión autóctona de fiebre por VZIK en la Isla de Pascua, no existía evidencia de la presencia del virus en el hemisferio occidental. Este brote fue limitado ya que la enfermedad figura en los registros de la isla hasta junio de 2014.
A finales de 2014 y comienzo del 2015 se dio un brote de una enfermedad exantemática, con varios miles de casos en la región nororiental de Brasil. Finalmente, el 7 de mayo 2015 se confirmó oficialmente el primer caso autóctono de VZIK en un paciente de Bahía (ProMED, 2015). Para finales del 2015 se había confirmado la transmisión autóctona del virus en la mayoría de los estados, con un número estimado de casos entre 400.000 y 1.300.000 (Hennessey et al. 2016).
Se han planteado varias hipótesis sobre como el VZIK fue introducido en Brasil. Se consideró primero la idea que el virus había sido introducido durante el campeonato mundial de futbol, desarrollados en varias ciudades de esta nación entre junio y julio del 2014 (Hennessey et al. 2016). Debido a la ausencia de participantes de la Polinesia, de donde las evidencias filogenéticas sugieren el origen del VZIK circulante en Brasil, se planteó como una probable vía de introducción, el campeonato mundial de canoa desarrollado en Rio de Janeiro en agosto de 1914, con la participación de atletas originarios de varios países de Polinesia, región endémica para el VZIK (Musso et al. 2015).
Una vez establecido en Brasil, el VZIK se extendió a otras naciones del continente. En octubre del 2015 se confirmó la presencia del VZIK en Colombia en el Distrito Bolívar y luego en otros estados, con un estimado para mayo del 2016 de cerca de 75.000 casos reportados por el Instituto Nacional de Salud (http://www.ins.gov.co/boletin-epidemiológico/ BoletinEpidemiologico/2016Boletinepidemiologico semana19).
En Venezuela la presencia del VZIK se reportó por primera vez en noviembre del 2015 (European Centre for Disease Prevention and Control, 2015). El diagnóstico se realizó en el laboratorio nacional de referencia, Instituto Nacional de Higiene “Rafael Rangel”. Cuatro muestras resultaron positivas para el VZIK por la reacción en cadena de polimerasa en tiempo real. Los casos, cuyas edades estuvieron en el rango de 40 a 55 años, fueron mujeres del área limítrofe con Brasil (World Health Organization, 2015). Para finales de enero 2016 las autoridades de salud de este país reportaban la ocurrencia de aproximadamente 5.000 casos. Utilizando modelaje estadístico y datos oficiales, se estimaron para esa fecha 400.000 infectados con el VZIK en Venezuela (Castro, Julio S, comunicación personal). Paralelamente epidemiólogos de una organización no gubernamental, la “Red para la Defensa Nacional de Epidemiología” han propuesto un estimado parecido al propuesto por Castro. Si estos valores son correctos, Venezuela tendría una tasa de infecciones per cápita mayor que Brasil. Esto no es sorprendente, puesto que los índices aédicos (porcentaje de hogares con presencia de mosquitos) de Venezuela son los más altos de la región: aproximadamente 20%, comparados con 5% en Brasil (http://www.healthmap.org/site/diseasedaily/article/have-we-underestimated-zika-outbreakvenezuela-21916).
Para el 19 de mayo del 2016, 39 países del continente, incluyendo América del Norte, Centro, Sur y el Caribe, habían reportado casos autóctonos de VZIK (PAHO 2016). Esta expansión explosiva de casos se debe principalmente a dos factores: la distribución cuasi continental del mosquito Aedes aegypti, el principal vector del VZIK, así como también de los virus del dengue y del Chikungunya, y la falta de inmunidad de la población.
Esta emergencia explosiva del VZIK, con el consiguiente alto número de individuos infectados en los brotes recientes en los países del Pacifico Sur y de las Américas, permitió detectar la asociación de la infección con infrecuentes complicaciones neurológicas severas: el SGB en adultos en la Polinesia Francesa (Cao-Lormau et al. 2016) y la microcefalia congénita en neonatos en Brasil (Centro Europeo de Prevención y Control de Enfermedades, 2016; Schuler-Faccii et al. 2015). Efectivamente en Julio del 2015, ya Brasil había informado un incremento de casos de desórdenes neurológicos, incluyendo casos del SGB, en los estados más afectados por la transmisión del VZIK. Lo mismo se observó en otras naciones con brotes importantes del VZIK, incluyendo Colombia, República Dominicana, El Salvador, Honduras, Surinam y Venezuela. En general el SGB fue detectado alrededor de tres meses después de la confirmación de la circulación del VZIK en estas regiones.
Desde octubre de 2015 Brasil había reportado un aumento considerable de casos de microcefalia en neonatos y planteado su posible asociación con la infección de mujeres embarazadas con el VZIK (Soares de Araujo et al. 2016).Estas observaciones fueron luego confirmadas en otros países, incluyendo la Polinesia Francesa, donde estudios retrospectivos de las epidemias 2013-2014, en noviembre 2015, también confirmaron la ocurrencia de diferentes malformaciones del sistema nervioso en neonatos (Cauchemez et al.2016). Estudios retrospectivos de la epidemia en esta región también confirmaron convincentemente la asociación entre VZIK y los casos del SGB (Cao-Lormeau et al. 2016).
En consecuencia, de estos reportes, la Organización Mundial de la salud (OMS) el 1 de febrero 2016 declaró que la asociación entre el grupo de casos de microcefalia y otros desórdenes neurológicos, reportados en Brasil, que seguían una casuística similar a la observada en la Polinesia Francesa, constituían una emergencia de salud pública, de preocupación mundial, movilizando a nivel internacional las medidas correspondientes de prevención y control.
Biología del Zika
El VZIK es transmitido por mosquitos principalmente por el Aedes aegypti, el mismo que transmite el Dengue y Chikungunya (Hayes, E.B. 2009, CDC 2016). La infección por el VZIK es asintomática en aproximadamente el 80 % de los casos (CDC 2016, Duffy et al. 2009) y cuando el virus causa enfermedad, los síntomas generalmente son leves y espontáneos. Otras vías de transmisión para el VZIK han sido reportadas como son la sexual, materno-fetal y a través de transfusiones sanguíneas.
Al igual que otros virus del género Flavivirus, el VZIK es un virus ARN envuelto con cápside icosaédrica y su genoma está codificado en una sola cadena (monocatenario) no segmentada, de polaridad positiva, de aproximadamente 10,8 Kb, limitado por dos regiones no codificantes (5’NCR y 3’NCR). La poliproteína codificada es traducida y co- y posttraduccionalmente procesada por proteasas virales y celulares en tres proteínas estructurales, la cápside (C), la premembrana/membrana (PrM) y la envoltura (E), y siete proteínas no estructurales NS1, NS2A, NS2B, NS3, NS4A, NS4B Y NS5 (Faye et al. 2014; Lindenbach et al. 2009; Kuno et al. 2007).
El mosquito al picar deposita el virus en la epidermis y la dermis del huésped. Es poco lo que se conoce del mecanismo exacto de replicación del VZIK, sin embargo, se ha reportado que los fibroblastos de la piel y queratinocitos epidérmicos, así como las células dendríticas inmaduras son altamente permisivas a la infección con VZIK (Hamel et al. 2015). La infección de los fibroblastos de la piel rápidamente resulta en la presencia de un alto número de copias de ARN y un incremento gradual con el tiempo de la producción de partículas virales, lo que indica la replicación activa en las células infectadas.
Figura 2: Filogenia del virus Zika (en azul) y otros flavivirus.
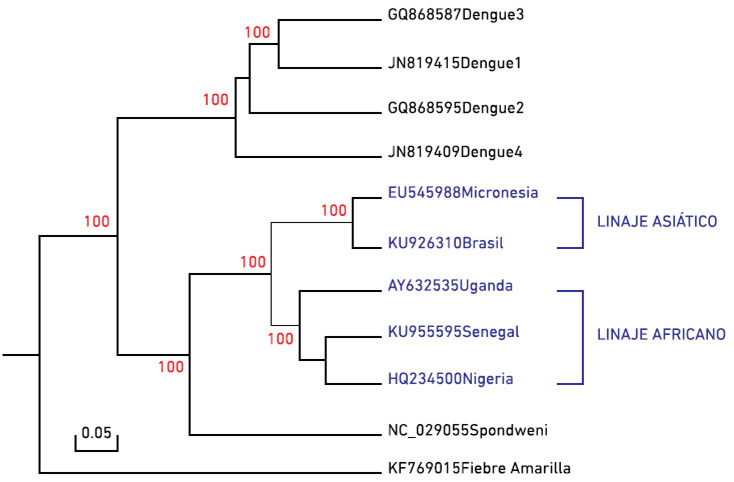
Se han descrito dos linajes del VZIK, el linaje africano y el linaje asiático. Algunos estudios filogenéticos indican que el virus que está expandiéndose por el continente americano está estrechamente relacionado con cepas provenientes del linaje asiático (Enfissi et al. 2016) (Figura 2), aunque el linaje africano oriental es el originario (Cao-Lormeau et al. 2014). El análisis de las secuencias disponibles del virus Zika por estudios filogenéticos de coalescencia sugieren un origen del virus hacia 1920 en Uganda (Faye et al. 2014).
Diagnóstico
La infección por el VZIK provoca un amplio rango de síntomas que son comunes con otras enfermedades transmitidas por Aedes tales como Dengue y Chikungunya, que hacen difícil el diagnóstico diferencial. El diagnóstico de la infección por el VZIK se basa en técnicas de aislamiento viral, detección de ARN y determinación de anticuerpos.
Durante la fase aguda de la enfermedad clínica (fase febril, virémica), se puede detectar el genoma viral en el suero de los paciente por técnicas de biología molecular tales como RT-PCR estándar o en tiempo real. Desde el día 5 de iniciados los síntomas, se puede detectar la respuesta serológica inducida por el virus (IgM y/o IgG). El ARN del VZIK también se ha detectado en la orina, semen y saliva de pacientes (Gourinat et al. 2015, Musso et al. 2015b, Musso et al. 2015c). En orina la deteccion del ARN se ha reportado hasta tres semanas después de iniciados los síntomas, en semen el virus puede permanecer por más tiempo que en la sangre, mientras que en la saliva la detección del genoma viral pareciera ser la misma que la observada en sangre. Estas observaciones sugieren que el diagnóstico molecular del VZIK, como ocurre para VD puede realizarse en orina después que ha desaparecido el virus en sangre, ampliando así la ventana para la detección del ARN del VZIK. Recientemente por RT-PCR también se ha identificado el ARN viral en fluído amniótico (Calvet et al. 2016; PAHO 2015), aunque la sensibilidad y especificidad del test para detectar la infección congénita es incierta.
No existen pruebas de diagnóstico serológico comercialmente disponibles o aprobadas por la Administración de Alimentos y Medicamentos (FDA) de los Estados Unidos para la detección de infección por el VZIK. Sin embargo, la FDA ha autorizado a laboratorios certificados durante la emergencia, el uso de dos herramientas de diagnóstico para el VZIK, el Zika MAC-ELISA para la detección cualitativa de anticuerpos IgM específicos en suero y fluido cerebroespinal. También ha autorizado el ensayo Trioplex Real-Time RT-PCR (Trioplex rRT-PCR), el cual es una prueba de laboratorio diseñada para detectar la infección por el VZIK, VD y VCHIK en humanos. Estas pruebas están siendo distribuidas a laboratorios certificadas para realizar pruebas en los Estados Unidos (http://www.cdc.gov/zika/hc-providers/ index.html).
La interpretación de los test serológicos es especialmente importante en el diagnóstico de VZIK. En una infección primaria con flavivirus existe poca reactividad cruzada con anticuerpos de otros virus genéticamente relacionados. Sin embargo, se ha demostrado que sueros de individuos con historia previa de infección con otro flavivirus (especialmente Dengue, fiebre Amarilla y Nilo del Oeste) pueden presentar reacción cruzada en las pruebas serológicas. En los ensayos de neutralización por reducción de placas (PRNT), los cuales ofrecen una mayor especificidad en la detección de anticuerpos neutralizantes (IgG), también se ha documentado la reacción cruzada.
En cuanto a los resultados para RT-PCR VZIK, un resultado negativo no descarta el diagnóstico de fiebre por VZIK debido a que el periodo de viremia es corto y variable. El periodo de incubación se ha descrito desde 3,5 días en humanos voluntarios (Bearcroft, 1956) hasta 6-10 días en viajeros retornando y donantes de sangre (Kutsuna et al. 2014; Musso et al. 2014).
Complicaciones del VZIK: comparación con VD y VCHIK
La patología del VZIK, así como de los VD y VCHIK, presenta un componente de inmunopatología muy marcado (Tabla 1).
Tabla 1. Complicaciones de VD, CHIK y VZIK
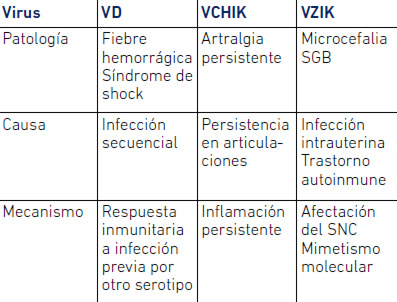
La inmunopatología que conduce a la manifestación hemorrágica de algunas infecciones secundarias por VD ha sido estudiada en extenso. Se sabe que el riesgo de la manifestación hemorrágica está relacionado con la infección por un segundo serotipo del VD. En esa manifestación participan anticuerpos facilitadores, anticuerpos producidos contra el serotipo de la infección previa de virus dengue, que en lugar de impedir la infección del nuevo serotipo infectante, le permiten acceder a la célula y a otros tipos celulares. A través de la interacción del virus con estos anticuerpos no neutralizantes, el virus logra ser internalizado en las células a través del reconocimiento de la porción Fc de los anticuerpos, logrando así una mayor y más variada infección de células. El proceso de estimulación de citoquinas que desencadena esta infección, que varía entre individuos de acuerdo a su inmunogenética, es lo que terminará determinando la severidad del ataque al endotelio y por ende la manifestación hemorrágica (Rothman, 2011).
Aunque la infección por el VCHIK es considerada menos severa que la infección por VD, pueden existir casos severos de la enfermedad generalmente asociados a co-morbilidades (Caglioti et al. 2013). La infección está asociada igualmente en un porcentaje significativo de los casos a secuelas de artralgia, debido a la persistencia del virus en las articulaciones y la subsecuente inflamación debido a la respuesta inmunitaria al virus (Petitdemange et al. 2015).
Recientes evidencias sugieren una asociación entre la infección materna por el VZIK y manifestaciones fetales adversas, como la microcefalia congénita (ECDC, 2015; Oliveira et al. 2016). Tang et al. (2016) han demostrado en pruebas experimentales, que el VZIK infecta eficazmente células progenitoras neurales humanas (hNPCs) derivadas de células madre pluripotentes inducidas. La asociación entre zika y microcefalia ha sido demostrada también en modelos animales (Malkhi, 2016).
También se asocia la infección por VZIK con el Síndrome de Guillain-Barré (SGB). El SGB consiste en una neuropatía periférica de tipo autoinmune, debida frecuentemente a la producción de anticuerpos anti-gangliósidos (Anaya et al. 2016). La abundante presencia de estos gangliósidos en la mielina conduce al daño en la mielina de nervios periféricos y por ende a la parálisis (Yuki et al. 2012).
Las manifestaciones patológicas de la infección por VZIK, superaron las expectativas que se tenían para una infección que se pensaba en buena proporción asintomática. Se ha planteado la posibilidad de que esto se deba a un fenómeno de facilitación de anticuerpos por la epidemia del VD existente en la región latinoamericana. Recientes evidencias sugieren que anticuerpos dirigidos contra el VD podrían facilitar la infección in vitro del VZIK (Paul et al. 2016). Sin embargo, esta información debe ser tomada con cautela, hasta que se demuestre este fenómeno en un modelo in vivo.
Por otra parte, estudios moleculares confirman lo discutido previamente sobre la relativa rápida evolución del VZIK. En particular, ciertos cambios en aminoácidos de la proteína preM podrían estar asociados a un incremento en neurovirulencia y mayor capacidad adaptativa (Wang et al. 2016).
Control de la enfermedad
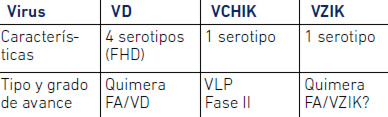
No existe hasta el presente una vacuna para VZIK, aunque algunas en desarrollo están basadas en las tecnologías utilizadas en las vacunas contra el dengue, que se basa en una quimera entre VD y Fiebre Amarilla (FA) (Tabla 2). Por otra parte, el uso de un anticuerpo monoclonal que reconoce todos los flavivirus (Dai et al. 2016) podría permitir desarrollar una eventual inmunoterapia contra el VZIK.
Tabla 2. Perspectivas de vacunas con virus dengue, chikungunya y Zika.
Por tanto las medidas de prevención son las mismas que se recomiendan para todas las enfermedades transmitidas por las picadas del mosquito Aedes que no tienen vacunas, que incluyen la protección individual a las picaduras de moquitos y el control del vector. Otro punto importante a considerar en el control de estas arbovirosis es la posibilidad de un reservorio selvático, es decir que primates no humanos puedan ser reservorios de la enfermedad. Se ha sugerido recientemente la infección de primates no humanos en Brasil (Favoretto et al. 2016), aunque se requieren más estudios para confirmar el real significado de estos hallazgos preliminares en la posibilidad de que se desarrolle un reservorio selvático en América.
La propagación del VZIK ha llevado a la necesidad de desarrollar nuevas estrategias de control de vectores dentro de las que destacan:
- Mosquitos genéticamente modificados, para los cuales la OMS ha recomendado mayores estudios de campo y la evaluación de riesgos para determinar el impacto de esta herramienta en la transmisión de enfermedades. Ensayos realizados han demostrado reducciones significativas en la población de Aedes Aegypti (Winskill et al. 2015).
- Liberación masiva de insectos machos previamente esterilizados mediante bajas dosis de radiación. Cuando los machos estériles se aparean los huevos de las hembras no son viables y la población de insectos disminuye (Bourtzis et al. 2016). Esta técnica de esterilización ya ha sido utilizada con éxito en el control de plagas (insectos) de importancia agrícola.
- Utilización de un método biológico de control, que utiliza la bacteria Wolbachia, la cual se encuentra en forma natural en el 60% de los insectos comunes (Johnson, 2015). Estas bacterias no infectan a los seres humanos u otros mamíferos, pero afectan la fertilidad de los mosquitos hembras, suprimiendo así las poblaciones de mosquitos. De esta manera los mosquitos no son modificados genéticamente, ya que la técnica no implica ninguna alteración genética. Además la bacteria bloquea en mosquitos la transmisión de varios patógenos humanos, incluyendo el VD y el VCHIK (Moreira et al., 2009), Se ha observado recientemente que la infección experimental de mosquitos con esta bacteria reduce totalmente su capacidad de transmitir el VZIK (Dutra et al. 2016).
Conclusiones
Un nuevo virus emergente azota la región latinoamericana, conduciendo a una situación bastante única, como lo es una triple epidemia de arbovirus (VD, VCHIK y VZIK). La epidemia por VZIK, inicialmente pensada en producir bajas secuelas desde un punto de vista de morbilidad, ha resultado un serio problema de Salud Pública, que ha encendido las alarmas para investigar más a fondo los mecanismos inmunopatológicos involucrados en esta infección, así como ha acelerado la necesidad de producir una vacuna contra este agente viral. El panorama nos sugiere que debemos prepararnos mejor para futuras epidemias, ya que existe una alta probabilidad que sigan emergiendo otras arbovirosis, como recientemente ha ocurrido de forma repetida en la región.
Tampoco podemos olvidar que existen otras epidemias muy activas en nuestro país, como la Malaria, que ha incrementado su extensión geográfica de afectación en el país, la fiebre hemorrágica venezolana que periódicamente resurge en el Estado Portuguesa y una epidemia de VIH que afecta a nuestra población Warao.
Referencias
- ALTHOUSE B.M., HANLEY K.A., DIALLO M., SALL A.A., BA Y., FAYE O., DIALLO D., WATTS D.M., WEAVER S.C., CUMMINGS D.A., (2015). “Impact of climate and mosquito vector abundance on sylvatic arbovirus circulation dynamics in Senegal”. Am J Trop Med Hyg 92: 88-97.
- ANAYA J.M., RAMIREZ-SANTANA C., SALGADOCASTANEDA I, CHANG C, ANSARI A., GERSHWIN M.E., (2016). “Zika virus and neurologic autoimmunity: the putative role of gangliosides”. BMC Med 14: 49-51
- BEARCROFT W.G., (1956). “Zika virus infection experimentally induced in a human volunteer”. Trans R Soc Trp Med Hyg 50: 442-448
- BOURTZIS K., LEES R.S., HENDRICHS J., VREYSEN M.J., (2016). “More than one rabbit out of the hat: Radiation, transgenic and symbiont-based approaches for sustainable management of mosquito and tsetse fly populations”. Acta Trop 157:115-30
- CAGLIOTI C., LALLE E., CASTILLETTI C., CARLETTI F., CAPOBIANCHI M.R., BORDI L., (2013). “Chikungunya virus infection: an overview”. New Microbiol 36: 211-227.
- CALVET G., AGUIAR R.S., MELO A.S., SAMPAIO S.A., DE FILIPPIS I., FABRI A., ARAUJO E.S., DE SEQUEIRA P.C., DE MENDONÇA M.C., DE OLIVEIRA L., TSCHOEKE D.A., SCHRAGO C.G., THOMPSON F.L., BRASIL P., DOS SANTOS F.B., NOGUEIRA R.M., TANURI A., DE FILIPPIS A.M., (2016). “Detection and sequencing of Zika virus from amniotic fluid of fetuses with microcephaly in Brazil: a case study”. Lancet Infect Dis: en prensa.
- CAO-LORMEAU VM., ROCHE C., TEISSIER A., ROBIN E., BERRY AL., MALLET HP., SALL AA., MUSSO D., (2014). “Zika virus, French polynesia, South pacific, 2013”. Emerg Infect Dis 20: 1085-1086.
- CAO-LORMEAU V.M., BLAKE A., MONS S., LASTÈRE S., ROCHE C., VANHOMWEGEN J., DUB T., BAUDOUIN L., TEISSIER A., LARRE P., VIAL A., DECAM C., CHOUMET V., HALSTEAD S., WILLISON H.J., MUSSET L., MANUGUERRA J.C., DESPRES P., FOURNIER E., MALLET H.P., MUSSO D., FONTANET A., NEIL J., GHAWCHÉ F., (2016). “Guillain-Barré Syndrome outbreak associated with Zika virus infection in French Polynesia: a case-control study”. Lancet 387: 1531–1539.
- CAUCHEMEZ, S., BESNARD,M., BOMPARD P., DUB P., GUILLEMETTE-ARTUR,P. , EYROLLE-GUIGNOT D., SALJE H., VAN KERKHOVE M., ABADIE V., GAREL C., FONTANET A., -MALLET P. H.(2016) Association between Zika virus and microcephaly in French Polynesia, 2013–15: a retrospective study. Lancet 387: 2125–2132.
- CDC. Zika virus. Atlanta, GA: US Department of Health and Human Services, CDC; 2016. http://www.cdc.gov/zika/index.html.
- CHADEE D.D., MARTINEZ R., (2016). “Aedes aegypti (L.) in Latin American and Caribbean region: With growing evidence for vector adaptation to climate change?”. Acta Trop 156: 137-143.
- CHOUIN-CARNEIRO T., VEGA-RUA A., VAZEILLE M., YEBAKIMA A., GIROD R., GOINDIN D., DUPONTROUZEYROL M., LOURENÇO-DE-OLIVEIRA R., FAILLOUX A.B., (2016). “Differential Susceptibilities of Aedes aegypti and Aedes albopictus from the Americas to Zika Virus”. PLoS Negl Trop Dis10: e0004543.
- DAI L., SONG J., LU X., DENG Y.Q., MUSYOKI A.M., CHENG H., ZHANG Y., YUAN Y., SONG H., HAYWOOD J., XIAO H., YAN J., SHI Y., QIN C.F., QI J., GAO G.F., (2016). “Structures of the Zika Virus Envelope Protein and Its Complex w`ith a Flavivirus Broadly Protective Antibody”. Cell Host Microbe 19: 696-704.
- DICK G.W., KITCHEN S.F., HADDOW A.J., (1952). “Zika virus. I. Isolations and serological specificity”. Trans R Soc Trop Med Hyg 46:509–520.
- DUFFY M.R., CHEN T.H., HANCOCK W.T., POWER A.M., KOLL J.L., LANCIOTTI R.S., PRETRICK M., MARFEL M., HOLZBAUER S., DUBRAY C., GUILLAUMOT L., GRIGGS A., BEL M., LAMBERT A.J., LAVEN J., KOSOY O., PANELLA A., BIGGERSTAFF B. J., FISHER M., HAYES E.B., (2009). “Zika virus outbreak on Yap Island, Federated States of Micronesia”. N Engl J Med 360: 2536-2543.
- DUTRA H.L., ROCHA M.N., DIAS F.B., MANSUR S.B., CARAGATA E.P., MOREIRA L.A., (2016). “Wolbachia Blocks Currently Circulating Zika Virus Isolates in Brazilian Aedes aegypti Mosquitoes”. Cell Host Microbe pii: S1931-3128: 30157-30163.
- ENFISSI A., CODRINGTON J., ROOSBLAD J., KAZANJI M., ROUSSET D., (2016). “Zika virus genome from the Americas”. Lancet 387: 227-228.
- EUROPEAN CENTRE FOR DISEASE PREVENTION AND CONTROL. (2015). “Zika virus epidemic in the Americas: potential associated with microcephaly and Guillain-Barré syndrome”. European Centre for Disease Prevention and Control, Stockholm, Sweden. http://ecdc.europa.eu/en/publications/publications/zika-virus-americanassociation-with-microcephaly-rapid-risk-assessment.pdf
- EUROPEAN CENTER FOR DISEASE PREVENTION AND CONTROL. 2016. Zika virus disease epidemic: potential association with microcephaly and Guillain-Barré syndrome (first update). European Centre for Disease Preventionand Control, Stockholm, Sweden. http://ecdc.europa.eu/en/publications /Publications/ rapid-risk-assessmentzika-virus-first-update-jan-2016.pdf
- FAVORETTO S., ARAUJO D., OLIVEIRA D., DUARTE N., MESQUITA F., ZANOTTO P., DURIGON E., (2016). “First detection of Zika virus in neotropical primates in Brazil: a possible new reservoir”. bioRXi.
- FAYE O., FREIRE C.C., IAMARINO A., FAYE O., DE OLIVEIRA J.V., DIALLO M., ZANOTTO P.M., SALL A.A., (2014). “Molecular evolution of Zika virus during its emergence in the 20(th) century”. PLoS Negl Trop Dis 8(1):e2636.
- GOURINAT A.C., O’CONNOR O., CALVEZ E., GOARANT C., DUPONT-ROUZEYROL M., (2015). “Detection of Zika virus in urine”. Emerg Infect Dis 21: 84 –86.
- HAMEL,R., DEJARNAC O., WICHIT S., EKCHARIYAWAT P., NEYRET A., LUPLERTLOP N., PERERA-LECOIN M., SURASOMBATPATTANA P., TALIGNANI L., THOMAS F., CAO-LORMEAU V-M., CHOUMET V., BRIANT L., DESPRÈS P., AMARA A., ISSEL H., MISSÉ D. J., (2015). “Biology of Zika Virus Infection in Human Skin Cells”. J Virol 89: 8880-8896
- HAYES E.B., (2009). “Zika virus outside Africa”. Emerg Infect Dis 15: 1347-1350. HENNESSEY M., FISCHER M., STAPLES J.E., (2016). “Zika virus spreads to new areas—region of the Americas”. MMWR Morb Mortal Wkly Rep 65: 55–58.
- JOHNSON, K., (2015). “The Impact of Wolbachia on Virus Infection in Mosquitoes”. Viruses 7: 5705–5717. KUNO G., CHANG G.J.J., (2007). “Full-length sequencing and genomic characterization of Bagaza, Kedougou, and Zika viruses”. Arch Virol 152: 687-696.
- KUTSUNA S., KATO Y., TAKASAKI T., MOI M.L., KOTAKI A., UEMERU H., MATONO T., FUJIYA Y., MAWATARI M., TAKESHITA N., HAYAMAWA S., KANAGAWA S., OHMAGARI N., (2014). “Two cases of Zika fever imported from French Polynesia to Japan, December 2013 to January 2014”[corrected]. Euro Surveill 19(4):pii20683.
- LEIS A.A., STOKIC D.S., (2012). “Neuromuscular manifestations of west nile virus infection”. Front Neurol 3: 37
- LEBRUN G., CHADDA K., REBOUX A.H., MARTINET O., GAÜZÈRE B.A., (2009). “Guillain-Barré syndrome after chikungunya infection ”. Emerg Infect Dis 15: 495–496.
- LINDENBACH B.D., RICE C.M., (2009). “Molecular biology of flaviviruses”. Adv Virus Res 59: 23-61. MACNAMARA F.N., (1954). “Zika virus: a report on three cases of human infection during an epidemic of jaundice in Nigeria”. Trans R Soc Trop Med Hyg 48:139–145.
- MALKKI H., (2016). “CNS infections: Mouse studies confirm the link between Zika virus infection and microcephaly”. Nat Rev Neurol: en prensa.
- MARCHETTE N.J., GARCIA R., RUDNICK A., (1969). “Isolation of Zika virus from Aedes aegypti mosquitoes in Malaysia”. Am J Trop Med Hyg 18: 411-415.
- MOREIRA, L.A., ITURBE-ORMAETXE, I., JEFFERY, J.A., LU, G., PYKE, A.T., HEDGES, L.M., ROCHA, B.C., HALL-MENDELIN, S., DAY, A., RIEGLER, M., ET AL. (2009). A Wolbachia symbiont in Aedes aegypti limits infection with dengue, Chikungunya, and Plasmodium. Cell 139, 1268–1278.
- MUSSO D., NHAN T., ROBIN E., ROCHE C., BIERLAIRE D., ZISOU K., SHAN YAN A., CAOLORMEAU V.M., BROULT J., (2014). “Potential for Zika virus transmission through blood transfusion demonstrated during an outbreak in French Polynesia, November 2013 to February 2014”. Euro Surveill 19: 1-3.
- MUSSO D., CAO-LORMEAU V.M., GUBLER D.J., (2015A). “Zika virus: following the path of dengue and chikungunya?”. Lancet 386: 243–244
- MUSSO D., ROCHE C., NHAN T.X., ROBIN E., TEISSIER A., CAO-LORMEAU V.M., (2015B). “Detection of Zika virus in saliva”. J Clin Virol 68: 53–55.
- MUSSO D., ROCHE C., ROBIN E., NHAN T., TEISSIER A., CAO-LORMEAU V.M., (2015C). “Potential sexual transmission of Zika virus”. Emerg Infect Dis 21: 359 –361.
- MUSSO D., GUBLER D.J., (2016). “Zika Virus”. Clin Microbiol Rev 29: 487-524.
- OLIVEIRA MELO A.S., MALINGER G., XIMENES R., SZEJNFELD P.O., ALVES SAMPAIO S., BISPO DE FILIPPIS A.M., (2016). “Zika virus intrauterine infection causes fetal brain abnormality and microcephaly: tip of the iceberg?. Ultrasound Obstet Gynecol 47: 6-7.
- PAHO, 2015. PAN AMERICAN HEALTH ORGANIZATION. Neurological syndrome, congenital malformations, and Zika virus infection. Implications for public health in the Americas—epidemiological alert. Washington DC: World Health Organization, Pan American Health Organization; 2015.
- PAHO 2016. PAN AMERICAN HEALTH ORGANIZATION / World Health Organization. Zika Epidemiological Update – 19 May 2016. Washington, D.C.: PAHO/WHO.
- PAUL L.M., CARLIN E.R., JENKINS M.M., TAN A.L., BARCELLONA C.M., NICHOLSON C.O., TRAUTMANN L., MICHAEL S.F., ISERN S., (2016). “Dengue Virus Antibodies Enhance Zika Virus Infection”. bioRxiv http://dx.doi.org/10.1101/050112
- PETITDEMANGE C., WAUQUIER N., VIEILLARD V., (2015). “Control of immunopathology during chikungunya virus infection”. J Allergy Clin Immunol 135: 846-855.
- PROMED-mail. 15 May 2015. Undiagnosed illness—Brazil (02): Zika virus confirmed. ProMED-mail archive no. 20150515.3364149. http://www.promedmail.org. Accessed 10 November 2015.
- PUJOL F.H., (2000): “Virus emergentes y reemergentes”. Vitae Nº3. http://caibco.ucv.ve/vitae/VitaeTres/homevitae.html.
- RAVI V., TALY A.B., SHANKAR S.K., SHENOY P.K., DESAI A., NAGARAJA D., GOURIE-DEVI M., CHANDRAMUKI A., (1994). “Association of Japanese encephalitis virus infection with Guillain-Barre syndrome in endemic areas of south India”. Acta Neurol Scand 90: 67–72.
- ROTHMAN A.L., (2011). “Immunity to dengue virus: a tale of original antigenic sin and tropical cytokine storms”. Nat Rev Immunol 11: 532-543.
- SCHULER-FACCINI L., RIBEIRO E.M., FEITOSA I.M.L., HOROVITZ D.D., CAVALCANTI D.P., PESSOA A., DORIQUI M.J., NERI J.I., NETO J.M., WANDERLEY H.Y., CERNACH M., EL-HUSNY A.S., PONE M.V., SERAO C.L., SANSEVERINO M.T., (2016). “Possible association between Zika virus infection and microcephaly—Brazil, 2015”. MMWR Morb Mortal Wkly Rep 65: 59–62.
- SOARES DE ARAÚJO, J.S., REGIS, CT, SILVA GOMES, R.G., TABARES, T.R., ROCHAS DOS SANTOS, C.R., MELO ASSUNCAO, P., VALERIA NOBREGA, R., ALVES PINTO, D.F., DANTAS BEZERRA, B.V., MATTOS, S.S. Microcephaly in northeast Brazil: a review of 16 208 births between 2012 and 2015. Bull World Health Organ. http://dx.doi.org/10.2471/BLT.16.170639
- SOLOMON T., DUNG N.M., VAUGHN D.W., KNEEN R., THAO L.T., RAENGSAKULRACH B., LOAN H.T., DAY N. P., FARRAR J., MYINT K.S., WARRELL M.J., JAMES W.S., NISALAK A., WHITE N.J., (2000). “Neurological manifestations of dengue infection”. Lancet 355: 1053–1059.
- SU S., WONG G., SHI W., LIU J., LAI A.C., ZHOU J., LIU W., BI Y., GAO G.F., (2016). “Epidemiology, Genetic Recombination, and Pathogenesis of Coronaviruses”. Trends Microbiol 24: 490-502.
- TANG, H., HAMMACK, C., OGDEN, S., WEN, Z., QIAN, X., LI., Y., YAO, B., SHIN, J., ZHANG, F., LEE, E., CHRISTIAN, K., DIDIER, R., JIN, P., SONG, H., MING, G. (2016). “Zika Virus Infects Human Cortical Neural Progenitors and Attenuates Their Growth”. Cell Stem Cell 18: 587-590
- TOLLE M.A., (2009). “Mosquito-borne diseases”. Curr Probl Pediatr Adolesc Health Care 39: 97-140.
- TROMBETTA C., PICCIRELLA S., PERINI D., KISTNER O., MONTOMOLI E., (2015). “Emerging Influenza Strains in the Last Two Decades: A Threat of a New Pandemic? Vaccines (Basel) 3: 172-185.
- WANG L., VALDERRAMOS S.G., WU A., OUYANG S., LI C., BRASIL P., BONALDO M., COATES T., NIELSEN-SAINES K., JIANG T., ALIYARI R., CHENG G., (2016). “From Mosquitos to Humans: Genetic Evolution of Zika Virus”. Cell Host Microbe 19: 561-565.
- WEAVER S.C., REISEN W.K., (2010). “Present and future arboviral threats”. Antiviral Res 85: 328-345.
- WINSKILL P., CARVALH D.O., CAPURRO M.L., ALPHEY L., DONNELLY C.A., MCKEMEY A.R., (2015). “Dispersal of Engineered Male Aedes aegypti Mosquitoes”. PLoS Negl Trop 9: e0004156.
- WORLD HEALTH ORGANIZATION. Disease Outbreak News: Zika virus infection – Venezuela [Internet]. 2015 [http://www.who.int/csr/don/03-december-2015-zika-venezuela/en/)
- YUKI N., HARTUNG H.P., (2012). “Guillain-Barré syndrome”. N Engl J Med 366: 2294-2304.
